|
Alpaca Info Library
What
is an Alpaca?
The
Alpaca (Vicugna pacos) is a
domesticated species of South American camelid
developed from the wild alpacas. It resembles a sheep in
appearance, but is larger and has a long erect neck as
well as coming in many colors, whereas sheep are
generally bred to be white.
Alpacas
are kept in herds that graze on the level heights of the
Andes of Ecuador, southern Peru, northern Bolivia, and
northern Chile at an altitude of 3500 to 5000 meters
above sea-level, throughout the year. Alpacas are
considerably smaller than llamas,
and unlike them are not used as beasts of burden but are
valued only for their fiber.
Alpacas only have fleece fibers, not woolen fibers, used
for making knitted and woven items much as sheep's wool
is. These items include: blankets, sweaters, hats,
gloves, scarves, a wide variety of textiles and ponchos
in South America, and sweaters, socks and coats in other
parts of the world. The fiber comes in more than 52
natural colors as classified in Peru, 12 as classified
in Australia and 22 as classified in America. Alpacas
and llamas differ in that llamas have banana shaped ears
and long tails and alpacas have straight ears and stubby
tails. Aside from these differences, llamas in general
are on average 1-2 feet taller, and bigger in proportion
than alpacas.
In
the textile industry, "alpaca" primarily
refers to the hair of Peruvian alpaca, but more broadly
it refers to a style of fabric originally made from
alpaca hair but now often made from similar fibers, such
as mohair, Icelandic sheep wool, or even high-quality
English wool. In trade, distinctions are made between
alpacas and the several styles of mohair and luster.
---------------------------------------------------------------------------------------------------------------------------------------

Color Inheritance by Gene Linkage
One theory of alpaca coat color inheritance stands apart from all the others. Researchers William L. Wall and Ron G. Cole, of Australia, who both own alpacas, propose that Mendel's rules of dominance and independent assortment do not entirely explain the inheritance of coat color in alpacas.
Wall's area of interest is agricultural sciences, especially genetics; Cole comes from a mathematics background. They propose a model of inheritance based on gene linkage.
The Wall and Cole theory of inheritance grew from their statistical analysis of matings that were registered by the Australian Alpaca Association's registry. In all, they studied the color of more than 10,000 cria from registered parents whose coat color was known. The results of these matings were compiled in two sets of coat color tables (presented in their entirety in tables 1-12): Version 1, which compiled the coat colors of over 7000
cria, and Version 2 which included the coat colors of an additional 3,000
cria.
Wall and Cole's theory of coat color inheritance in alpacas formed as result of analyzing Version 1 of the tables. They then used their theory to predict the color distribution among the additional
cria. These are the figures charted in Version 2. The accuracy of their predictions lends considerable credibility to their ideas.
The goal of the Wall and Cole research was to: 1. Determine the minimum number of genes necessary to explain the range of colors found in alpacas;
2. Map the genes on the chromosomes;
3. Explain the action of modifier genes;
4. Explain the action of the multi gene.
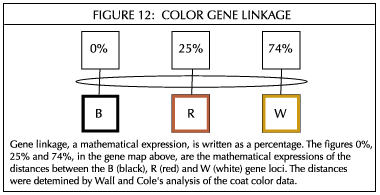
In the process, they concluded that coat color inheritance was determined by the process of gene linkage and not by dominance and simple assortment. They further concluded that there were five genes total: three primary color genes - black, red, and white - which are linked on the same chromosome; a modifier gene which determines the amount of color; and a multi gene which determines the distribution of color. Walland Cole hypothesize that the chromosomes carrying the three linked color genes resemble the above diagram.
Once Wall and Cole settled on the gene linkage method of inheritance, and determined from their coat color tables the relative distance apart of the linked genes, they were ready to predict the outcome of the additional matings that were included in Version 2 of the coat color tables. Their predictions were more than 90 percent accurate.
Because the B, R, and W genes are linked, this allows for 64 possible genotypes (4 alleles X 4 alleles X 4 alleles = 64) which are expressed as 27 phenotypes. This conclusion is reached by taking the B (black) gene, the alleles of which are B and b (where BB, Bb,
bB, or bb represent four possibilities), and making the same assumption for R and W, therefore 4 X 4 X 4 = 64. However, as Bb and bB are indistinguishable, there are three phenotypes (BB, Bb, and bb). The same is true for R and W. Therefore3 X 3 X 3 = 27 phenotypes.
In similar fashion, Wall and Cole theorized that the diluter gene has four genotypes and three phenotypes: DD,
Dd, dD, and dd. When you take the 27 color phenotypes available and multiply them by the three diluter gene phenotypes, the result is a potential for 81 different phenotypes. This range of possible color shades explains every conceivable alpaca color. These colors would occur on a continuous variation from light to dark, red to brown, fawn to white, etc.
The research derived from the color tables also led Wall and Cole to theorize that there are three alleles of the multi gene: O, o, and ø with solid (O) dominant. The multicolored coat in alpacas is expressed in many forms. These forms include
1. A small white blaze on the face of an otherwise totally black animal;
2. Boots (i.e., feet and lower leg colors different from the coat color expressed over the rest of the animal);
3. White on white or black on black (i.e., white spots on a white-coated animal or black patches on a black-coated animal which, because of the base color of the animal's coat, are unseen as spots or patches).
All grays in this genetic context are considered multis, with the possible exception of "true solid gray."
Calculating the various possible phenotypes that would occur from specific matings under this theory establishes that a two-to-one ratio of solid to multicolored animals would result from matings of multicolored parents. This conclusion is also consistent with the data found in the tables. Finally, their research confirmed that all grays were multis with the black, red, and white genes operating.
Wall and Cole's research was verified independently by examining published data presented by Rigoberto Calle Escobar who, in his book Animal Breeding and Production of American
Camelid, reported the following results of a color mating study conducted at La Raya Ranch:
From observations made at La Raya Ranch, 1,000 white females mated with white sires produced 50 to 60 percent white offspring; 19 percent were light fawn; 17 percent were patched. In decreasing order came cinnamon, light coffee, dark coffee and black. It was also verified that from every 300 offspring of the white with white cross, only one completely black offspring was produced. Similarly from the crossing of white sires with other colored females (with exception of light spotted fawn) a predominance of the mothers' color was noted. In the case of females with light fawn and spotted, 40 percent of the offspring were white. These results of color crosses which have been verified, reinforce the thesis that color inheritance is complex and is based on many pairs of genes which, because of a not very intense selection in the herds, are maintained in a pool of genes of the population, conserving color variability.
It is interesting to note how Wall and Cole's study's predictive value holds up in explaining the results of the La Raya color mating study. Escobar's La Raya observations and Wall and Cole's calculations from the Australian herd when white was mated to white follows in Figure 12.

Breeding for Color in Practice
What happens as a practical matter when you breed white to white, black to black, one color to a different color or solid color to multicolor? Alpaca breeders are fortunate to have Wall and Cole's exhaustive study of coat color inheritance which is intended to be an easy reference for breeders (see Tables 1-12). The study is based on the phenotypic color of the parents and their progeny; it is not intended to suggest the alpaca's genotype.
The base data for the Wall and Cole work was derived from the Australian Alpaca Association registration database which records alpaca registrations with designated colors. The tables were created from registrations as of March 1996 and included 10,849 alpacas.
There are two types of tables:
1. The solid color cross table, which presents the progeny from crosses of sires and dams of the same color. Numbers of crosses and sex of progeny are listed, together with numbers of cria for each solid and each multiple color registered (Tables 1-8).
2. The multi or grey color cross tables which list number of matings and sex of progeny, together with results of analysis of each color of male crossed with each color of female and vice versa for each of the colors. There are four of these tables (Tables 9-12).
It should be understood that the color tables can not be used to predict the outcome of a specific cross between two animals. The data presented is an analysis of the combination of all available data. It is meant to present the results of past experience. An alpaca breeder might choose to study the various tables to determine what has transpired in the Australian National Herd as a guide to the likelihood of various possible color outcomes from specific
breedings. Wall and Cole suggest that readers of their coat color tables pay attention to the "white space" in the tables. They point out that the absence of offspring of particular colors, as evidenced by "white space," is as informative as the offspring recorded in the tables.
Observations On Color Matings
In the Australian color mating tables (Tables 1-12), the color of the alpacas were grouped as follows:
1. Fawn and roan alpacas were assigned to red;
2. Silver grays and blacks were assigned to black;
3. Browns were assigned to brown;
4. Whites were assigned to white;
5. Multicoloreds were assigned according to the mix of colors listed, for example, a dark fawn/light fawn/white alpaca was assigned to red; a dark fawn/medium gray alpaca (roan) was assigned to brown.
Understanding this, you can use the charts to make the following observations: 1. When breeding white to white, the progeny were 60 percent white; 18 percent red; 17 percent brown; and five percent black.
2. When breeding white to brown, the progeny were 43 percent brown; 10 percent black; 27 percent red; and 20 percent white.
3. When breeding black to black, the progeny were 85 percent black; 11 percent brown; one percent red; and three percent white.
4. When breeding white to black the progeny were 24 percent white; 14 percent red; 30 percent black; and 32 percent brown.
5. When breeding brown to black the progeny were 52 percent brown; 40 percent black; three percent red; and five percent white.
The Bottom Line
The color of the progeny can often be predicted with accuracy if the breeder is familiar with the stud being used, particularly if he has sired a large number of offspring. A famous alpaca stud, Hemingway, is a good example. He has been bred to more than 30 black females. All the offspring, 100 percent, have been fawn, mostly dark fawn. When Hemingway is bred to solid-colored females, such as brown or fawn, he almost always produces a lighter colored cria in the same basic color of the mother; when bred to white females, he produces white
cria. Accoyo's El Moustachio (white) and Accoyo's Victor (fawn) often produce a cria the color of the mother, especially Victor, who has thrown a lot of black cria when mated to black females.
The highest likelihood for creating a certain color occurs when mating two alpacas of the same color. Alpacas seem to carry a variety of color genes, especially white alpacas. If Cole and Wall are correct, every alpaca carries every color. When crossing a white alpaca with a colored alpaca, the progeny are more likely to be colored than white by a considerable margin. Two colored alpacas almost always result in colored progeny. Pintos can pop up almost anywhere or, as Barreda says, "pintos are hard to get rid of."
Alpaca breeders need to form their own goals as to colors. If they want to produce unique colors for the pet market, they can mix up solids with
multicolors, black with white, and so on. If their goals involve eventually producing commercially valuable fiber, they can breed solid to solid, preferably white.
|












